|
Leffingwell &
Associates
|
Translate |
John C. Leffingwell,
Ph.D.
Leffingwell &
Associates
|
Leffingwell &
Associates
|
Translate |
John C. Leffingwell,
Ph.D.
Leffingwell &
Associates
The sense of smell is a primal sense for humans as well as animals. From an evolutionary standpoint it is one of the most ancient of senses. Smell (or Olfaction) allows vertebrates and other organisms with olfactory receptors to identify food, mates, predators, and provides both sensual pleasure (the odor of flowers and perfume) as well as warnings of danger (e.g., spoiled food, chemical dangers). For both humans and animals, it is one of the important means by which our environment communicates with us.
This paper will explore the current status our understanding of olfaction and provide in some detail the possible molecular interactions that specify odorant signaling.
General Physiology of Olfaction
Odorants are volatile chemical compounds that are carried by inhaled air to the Regio olfactoria (olfactory epithelium) located in the roof of the two nasal cavities of the human nose, just below and between the eyes.
The odorant must possess certain molecular properties in order to provide sensory properties. It must have some water solubility, a sufficiently high vapor pressure, low polarity, some ability to dissolve in fat (lipophilicity), and surface activity. And to date, no known odorant possesses a molecular weight greater than 294.1
The olfactory sense is able to distinguish among a practically infinite number of chemical compounds at very low concentrations.2
The olfactory region of each of the two nasal passages in humans is a small area of about 2.5 square centimeters containing in total approximately 50 million primary sensory receptor cells.
The olfactory region consists of cilia projecting down out of the olfactory epithelium into a layer of mucous which is about 60 microns thick. This mucous layer is a lipid-rich secretion that bathes the surface of the receptors at the epithelium surface. The mucous layer is produced by the Bowman’s glands which reside in the olfactory epithelium. The mucous lipids assist in transporting the odorant molecules as only volatile materials that are soluble in the mucous can interact with the olfactory receptors and produce the signals that our brain interprets as odor. Each olfactory receptor neuron has 8-20 cilia that are whip-like extensions 30-200 microns in length. The olfactory cilia are the sites where molecular reception with the odorant occurs and sensory transduction (i.e., transmission) starts.
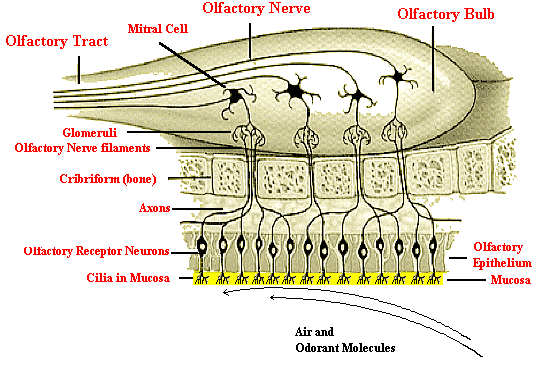
Above the mucous layer is the base olfactory epithelium which consists partially of basal cells located in the lowest cellular layer of the olfactory epithelium which are capable of mitotic cell division to form olfactory receptor neurons when functionally mature. The olfactory receptor neurons turnover approximately every 40 days. The epithelium also contains pigmented cells that are light yellow in humans and dark yellow to brown in dogs. The depth of color seems to be correlated with olfactory sensitivity.
While the olfactory receptor neurons extend through the epithelium to contact odorants in the atmosphere, on the opposite side within the epithelium, the neuronal cells form axons that are bundled in groups of 10-100 to penetrate the ethmoidal cribiform plate of bone, reaching the olfactory bulb of the brain where they converge to terminate with post-synaptic cells to form synaptic structures called glomeruli. The glomeruli are connected in groups that converge into mitral cells. (Note that in the picture above this convergence is not clearly depicted). For example, in rabbits, there are 26,000 receptor neurons converging onto 200 glomeruli which then converge at 25:1 onto each mitral cell. The total convergence is estimated to be about 1000:1.3
Physiologically, this convergence increases the sensitivity of the olfactory signal sent to the brain. From the mitral cells the message is sent directly to the higher levels of the central nervous system in the corticomedial amygdala portion of the brain (via the olfactory nerve tract) where the signaling process is decoded and olfactory interpretation and response occurs.
The Trigeminal Sense in the Olfactory Epithelium
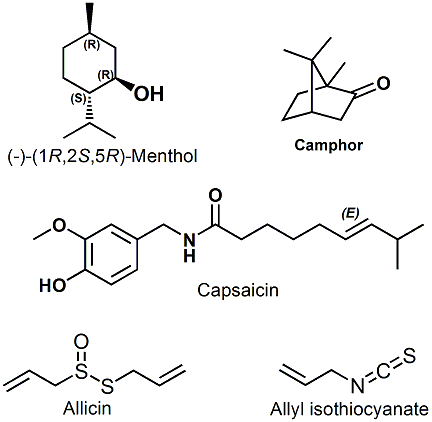
It must also be recognized that the olfactory epithelium contains another sensory system in the form of "Trigeminal Nerve" receptors. Whereas, the olfactory receptor system is highly localized in humans, the fifth cranial or trigeminal nerve (which is the largest cranial nerve and is the responsible sensory nerve of the face, teeth, mouth, most of the scalp, and the motor nerve of the muscles of mastication) provides a second set of nerve endings which are responsible for tactile, pressure, pain and temperature sensations in the areas of the mouth, eyes and nasal cavity. A number of chemical trigeminal stimulants produce effects described as hot, cold, tingling or irritating. For example, "leavo-menthol or (-)-menthol" produces the trigeminal feeling of cold at moderate concentrations and "hot" at high concentrations in the nasal cavity. This type of sensory "description" is often not just limited to the areas of the nose, mouth and eyes, but also occurs on skin areas not served by the 5th cranial nerve (especially, the genitalia) and thus such stimulants may effect a variety of nerve endings. Similarly "camphor" which possesses markedly more aroma than menthol, also produces the "cold" sensation via interaction with trigeminal receptors. Ohloff states that "About 70% of all odors are said to stimulate the trigemenal nerve although, in general, they may be several times less sensitive than olfactory receptors".4
Other commonly encountered trigeminal stimulants include the chemicals allyl isothiocyanate (mustard, mustard oil), capsiacin (hot Chile powder, mace spray) and Allicin (garlic).
|
|
|
|
|
|
Trigeminal neurons mediate the detection of thermal, mechanical, and chemical stimuli. For example, chemosensory innervation of the nasal epithelium utilizes two major trigeminal fiber systems - unmyelinated C-fibers and myelinated Ad-fibers. C-fibers are involved in the burning pain sensation whereas Ad-fibers mediate sharp, stinging sensations.4c
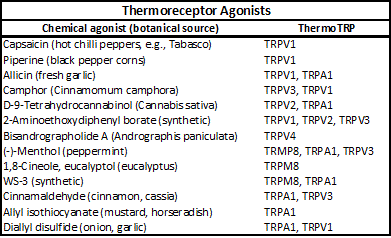
In the case of thermoreceptors, these are activated when a thermal (or chemical) stimulus excites primary afferent sensory neurons of the dorsal or trigeminal ganglia. In the last 20 years, there has been tremendous progress in determining the various receptor structural sequences. Thermoreceptors belong to the class of transient receptor potential (TRP) channels of which seven subfamilies exist (TRPC, TRPV, TRPM, TRPA, TRPP, TRPML, and TRPN). Six members of three TRP subfamilies are involved in mammalian temperature-sensitive thermoreception4a.
The closely related TRPV analogs are activated by heat, TRPV1 (=/>43oC), TRPV2 (=/>52C), TRPV3 (22–40oC), and TRPV4 (>~27oC), while TRPM8 (<~28oC) and TRPA1 (<~18oC) are activated by cold4a.
Certain types of chemical agonists activate these same thermoTRP channels. TRPV1 was the first thermoreceptor characterized and is referred to as a vanilloid receptor, as it is activated by capsaicin as well as heat. The cold and menthol receptor, TRPM8, was characterized by McKemy, Neuhausser, and Julius4d and by Peier et al.4e in 2002.
Paradoxically, the cold receptor TRPA1, which is activated by noxious cold to produce a pain-like sensation, produces a human sensorial effect often described as “hot.”
The table above, entitled "Thermoreceptor Agonists" provides examples of chemical agonists that activate these thermoTRPs.
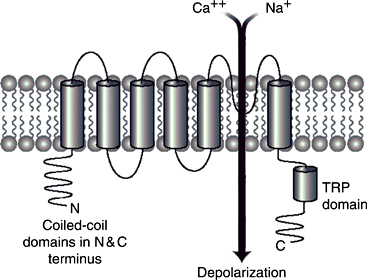
All of these thermoTRPs are gated Ca++ channels consisting of six transmembrane domains (TM1–TM6) flanked by large N- and C-terminal cytoplasmic domains4a. A schematic representation is shown in the figure above, labeled "TRPM8 receptor channel", with the putative ion channel between TM5–TM6 in TRPM8, which is activated by menthol and other cold stimuli.
For a recent review of other peripheral factors that modulate olfaction see the review by Mary Lucero4f.
_____________
1. Demole, E., H. Wuest, Synthèses stéréosélectives de deux trioxydes C18H30O3 stéréoisomères, d’ambréinolide et sclaréol-lactone a partir de derives du (+)-manool, Helv. Chem. Acta, 50: 1314 (1967).
2. Ohloff, G., Scent and Fragrances, Springer-Verlag, Berlin Heidelberg, 1994
3. http://www.cf.ac.uk/biosi/staffinfo/jacob/teaching/sensory/Copy%20of%20olfact1.html
4. Ohloff, G., Scent and Fragrances, Springer-Verlag, Berlin Heidelberg, 1994, p. 6
4a. Leffingwell, J.C., Cooling Ingredients and Their Mechanism of Action, Chapter 65, Handbook of Cosmetic Science and Technology, 3rd ed., Andre O. Barel, Marc Paye, Howard I. Maibach, Eds., Informa Healthcare (Pub.), New York, 2009. p. 670-671; See also Voets, T., Talavera, K., Owsianik, G. and Nilius, B., 2005. Sensing with TRP channels. Nature chemical biology, 1(2), pp.85-92.
4b. The figure for the "TRPM8 receptor channel" is from the 2009 Leffingwell reference 4a. This exact figure, without any citation or acknowledgement, is also found in Bharate, S.S. and Bharate, S.B., 2012. Modulation of thermoreceptor TRPM8 by cooling compounds. ACS chemical neuroscience, 3(4), pp.248-267.
4c. Meusel, T., Negoias, S., Scheibe, M. and Hummel, T., 2010. Topographical differences in distribution and responsiveness of trigeminal sensitivity within the human nasal mucosa. PAIN®, 151(2), pp.516-521.
4d. McKemy DD, Neuhausser WM, Julius D. Identification of a cold receptor reveals a general role for TRP channels in thermosensation. Nature 2002; 416:52–58.
4e. Peier AM, Moqrich A, Hergarden AC, et al. A TRP channel that senses cold stimuli and menthol. Cell 2002; 108(5):705–715.
4f. Lucero, M.T., 2013, January. Peripheral modulation of smell: fact or fiction?. In Seminars in cell & developmental biology (Vol. 24, No. 1, pp. 58-70). Academic Press.
_____________
![]() Email:
Email:
leffingwell@mindspring.com
 |
 |
|
|
|
|